ЭМОЦИОНАЛЬНАЯ ПАМЯТЬ ПРИ ФОРМИРОВАНИИ ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ ЖИВОТНЫХ ДО И ПОСЛЕ НАСЫЩЕНИЯ
ЭМОЦИОНАЛЬНАЯ ПАМЯТЬ ПРИ ФОРМИРОВАНИИ ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ ЖИВОТНЫХ ДО И ПОСЛЕ НАСЫЩЕНИЯ

Аннотация
Выявленные закономерности последовательного выбора животными целей поведения до и после насыщения связаны с фиксацией в эмоциональной памяти эмоций, возникающих при достижении результата. Введено понятие «эмоционально-памятных мотиваций», направляющих поведение на достижение и избегание зафиксированных в эмоциональной памяти положительных и отрицательных переживаний. Установлена природа «холостых реакций», возникающих после насыщения в виде отказа от достигаемых результатов, направляемых не потребностью, а стремлением получать закреплённые в эмоциональной памяти положительные эмоции. Предложена модификация функциональной системы с учетом эмоциональной памяти и структуры мотиваций целенаправленного поведения.
1. Введение
В настоящее время одним из наиболее важных вопросов в проблеме целенаправленной деятельности человека и животных являются особенности её изменения в процессе удовлетворения потребности , , , . Большое внимание при этом уделяется изучению эмоций и их влиянию на формирование целей поведения. На эту тему существует ряд теорий.
Известна «Биологическая теория эмоций» П.К. Анохина , о роли эмоций в системной организации целенаправленного поведения. К.В. Судаковым определены индивидуальные изменения вегетативных функций при однотипных эмоциональных нагрузках в системных механизмах эмоционального стресса , . «Информационная теория эмоций» П. В. Симонова определяет зависимость эмоций от потребности с учетом разности между необходимой и реально существующей информацией. Теория «Эмоциональных мотиваций» Е.А. Пырьева рассматривает мотивационные функции эмоций , , . «Динамическая теория эмоций» Е.А. Юматова , , , посвящена вопросам последовательного развития эмоциональных состояний в процессе целенаправленного поведения с учетом исходного прогноза вероятности и реального достижения результата.
В этих представлениях речь идет о реально действующих эмоциях, принимающих участие в афферентном синтезе и определяющих, совместно с доминирующей мотивацией, общую направленность целенаправленного поведения. Вместе с тем, «общая направленность» не определяет принятие решений в ситуации последовательного выбора одного из нескольких вариантов достижения положительного результата. Для «выбора» необходима модель о ранее испытанных эмоциональных переживаниях для каждого из поведенческих актов — так называемая «аффективная» или «эмоциональная память» , — запоминание, хранение и воспроизведение связанных с эмоциями событий (в основном связана с миндалевидным телом, которое отвечает за обработку эмоций и их запоминание). Согласно этим исследованиям, эмоциональная память может быть стимулом, вызывающим или тормозящим действие, в зависимости от активируемых в памяти эмоциональных переживаний.
Эта память, как и память в целом, существенно связана с изменением доминирующей мотивации. Поэтому задача исследования состоит в экспериментально-теоретической проверке гипотезы об участии эмоциональной памяти в механизмах принятия решении при последовательном выборе целенаправленных актов на разных стадиях насыщения.
2. Методы и принципы исследования
Опыты приведены на крысах линии Wistar (самцы весом 150–180 г.) при двухсуточный водной депривации. Жажду животные удовлетворяли при потреблении ими порций воды (по 0,5 мл) в 2-х лунках лабиринта.
2.1. Аппаратная часть опытов
Схема лабиринта реализует ситуацию последовательного выбора животными одного из двух вариантов достижения результата (рис. 1). В его основе лежит схема простого симметричного Т-образного лабиринта, дополнительно снабженного возвратными коридорами и дверками, определяющими направления перемещения животных.
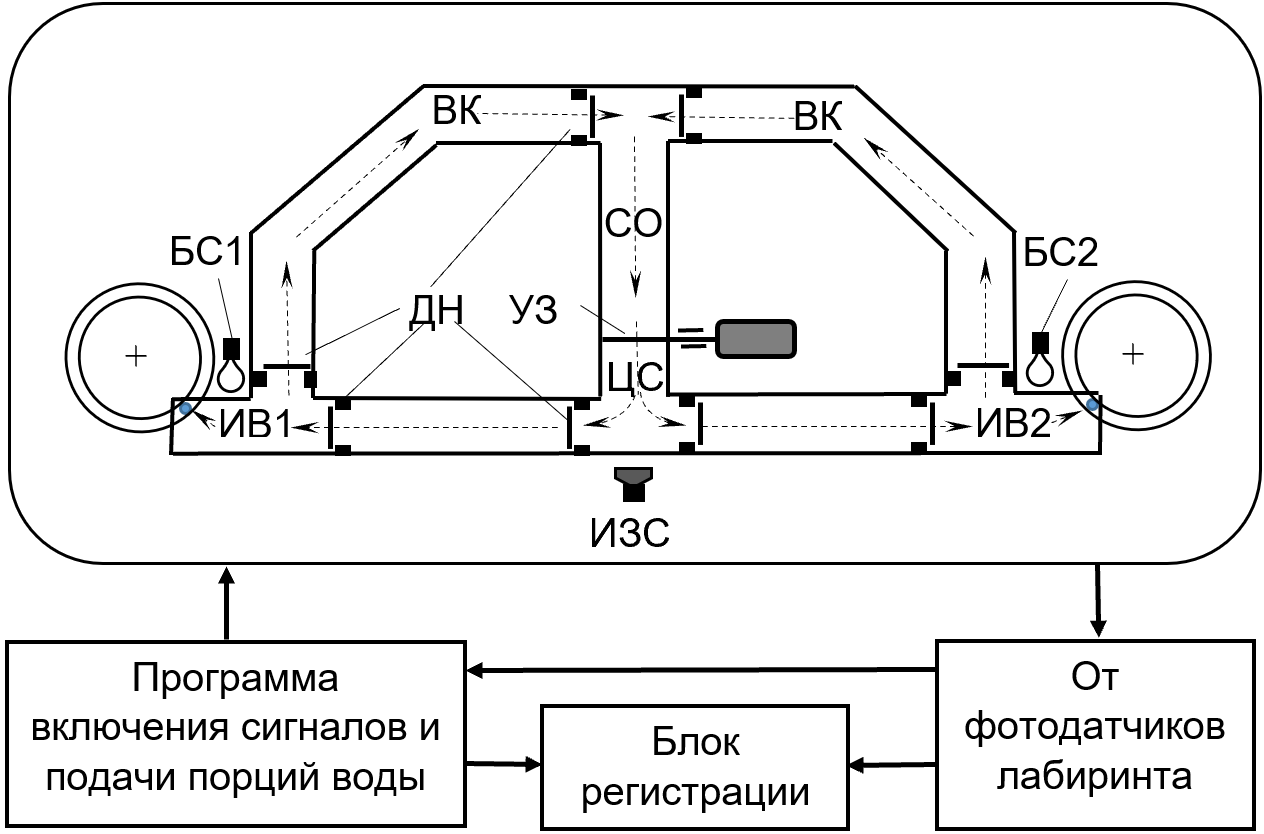
Рисунок 1 - План Т-образного лабиринта с возвратными коридорами и дверками направлений перемещения животных
Звуковые и световые сигналы выключались при попадании животного к источникам воды.
Для перемещения крыс в каждом поведенческом акте только к одному из ИВ и возврату в СО (стрелки с пунктирной линией) предусмотрены дверки направления (ДН). ИВ выполнены в виде лунок, расположенных по окружности дисков по бокам лабиринта. При попадании животного к одному из ИВ, в соответствии с программой опыта, диск мог осуществить, или не осуществить вращение. В первом случае в выбранном ИВ появлялась лунка с водой. Во втором случае животное получало пустую лунку.
Управление опытами проводилось устройством с «Программой включения сигналов и подачи порций воды», работающим благодаря сигналам «От фотодатчиков лабиринта». Фиксация поведенческих актов, включаемых сигналов и подачи порций воды осуществлялась «Блоком регистрации».
Применение лабиринта позволило представить целенаправленную поведенческую активность животного в виде дискретной последовательности чётко очерченных поведенческих актов с их результатами, что являлось несомненным упрощением модели поведения в ситуации выбора даже по отношению к той, которая реализуется в простом Т-образном лабиринте без ВК и без ДН. Но именно благодаря такому упрощению и стал принципиально возможным анализ последовательного выбора животными целей поведения.
Приведённый методический подход соответствует представлениям К. В. Судакова о «системном квантовании поведенческой деятельности», согласно которому вся деятельность представляет собой непрерывную смену последовательных «квантов поведения» , .
2.2. Подготовка крыс для проведения основных опытов
Перед основными исследованиями с животными проводились 7 опытов «предварительной выработки навыка» (ПВН): порции воды они получали в том ИВ, со стороны которого подавался световой боковой сигнал (БС). Чтобы в процессе ПВН животные не смогли усвоить никакой закономерности в подаче БС1 и БС2, они включались «чисто случайно» и равновероятно.
В процессе ПВН завершался «системогенез» обоих целенаправленных актов — побежек из ЦС к ИВ1 и к ИВ2, то есть становление и автоматизация системной архитектуры двух однотипных «квантов» поведения , .
2.3. Процедурная часть исследования
По окончании ПВН с животными проведен ряд экспериментальных исследований на различных стадиях насыщения в условиях «Направляемого» и «Свободного выбора» источников воды. При «Направляемом выборе» подавались световые боковые сигналы, указывающие на сторону подачи порции воды. В отличие от этого, в опытах «Свободного выбора» этот сигнал не подавался, и животные принимали решения без получения направляющих сигналов. В этом условии проведены 2 эксперимента — при двусторонних отрицательных результатах и при переделке стороны получения порции воды.
Статистический анализ данных осуществлялся пакетом Microsoft Excel. Определялись средние по совокупности опытов частоты выбора источников воды (%) ± ошибка среднего.
Достоверности различий между частотами определялись по статистическим критериям К. Пирсона Хи-квадрат и φ-Фишера , , .
3. Экспериментальное исследование
3.1. «Направляемый выбор» источников воды до и после насыщения
В 1-м эксперименте животные получали воду только в том ИВ, со стороны которого включался БС (аналогично ранее проведенной процедуре ПВН, т.е. равновероятно слева и справа). Каждый опыт прекращали после того, как животные полностью прекращали побежки и засыпали.
Исследование проведено на 10 крысах по 10–15 опытов с каждой. Определяли число ошибочных побежек, когда животные выбирали ИВ, противоположный включенному боковому сигналу.
В процессе опыта характер потребления крысами порций воды изменялся постепенно. До насыщения животные полностью потребляли воду, вылизывая питьевые лунки насухо. На средней стадии насыщения в лунках появлялись постепенно возрастающие остатки воды. После насыщения крысы полностью отказывались от порций воды, но при этом продолжали совершать побежки к ИВ — так называемые «холостые реакции» (см., например, ).
Сохранился единый элемент всех побежек — их конечный этап: при попадании в отсек с источником воды животные неизменно устремлялись к водной лунке. Именно начиная с этого момента наблюдалось отличие в их поведении. До насыщения животные насухо вылизывали лунку и быстро возвращались в СО. После насыщения они отскакивали от питьевой лунки, оставив порцию не выпитой, проявляли характерное фырканье и резкие движения. Быстро отскочив от ИВ, они медленнее возвращались в СО. После 20–30 «холостых реакций» животные постепенно их прекращали и засыпали.
В среднем по всем опытам получены общие частоты ошибок по всем поведенческим актам, а также частоты ошибок при повторном включении бокового сигнала с одной стороны и частоты ошибок при смене стороны включения бокового сигнала, определенные «До» и «После» насыщения.
Общая частота ошибок до насыщения составила 4,5±0,4%, а после насыщения она достоверно возросла до 16,8±0,7% (P<0,001).
До насыщения частота ошибок при повторном включении БС с одной стороны составила 3,2±0,5%; а при смене стороны включения БС она оказалась почти в два раза выше: 5,9±0,6%. Различие этих частот достоверно (p<0,01).
Т.е. ошибки чаще появлялись при включении БС на стороне, противоположной стороне предыдущего получения порции воды: животные совершали ошибку, предпочитая выбирать тот ИВ, где перед этим была получена порция воды.
По-видимому, эти «неправильные» реакции связаны не с ошибочной побежкой на боковой сигнал, а со стремлением достигнуть тот ИВ, который перед этим завершился положительным результатом и, согласно биологической теории эмоций П.К. Анохина, вызвал положительную эмоцию , . Согласно этой теории, положительное эмоциональное состояние при удовлетворении потребности возникает в том случае, если обратная информация от результатов выполненного действия отражает все компоненты положительного результата и совпадает с акцептором действия. Побежки против бокового сигнала при наличии потребности, скорее, не являются «ошибками» — это целенаправленные поведенческие акты для повторного достижения испытанной в предыдущем акте положительной эмоции. Т.е. животные, игнорируя боковой сигнал, предпочитали выбирать тот ИВ, где перед этим была получена порция воды и испытана положительная эмоция.
Иной характер ошибок проявился на стадии «после насыщения». Частота ошибок при повторном включении БС с одной стороны составила 29,9±1,3%; тогда как при смене стороны включения БС она оказалась равной 3,7±0,5% с высокой достоверностью их различия (P < 0,001). Видно, что после насыщения частоты ошибок при повторном включении БС на одной стороне лабиринта существенно возросли (3,2±0,5% и 29,9±1,3%; P < 0,001).
В отличие от полноценной потребности, после насыщения животные намного чаще совершали ошибки при повторном включении БС на одной и той же стороне лабиринта, предпочитая не выбирать тот ИВ, где перед этим они получили порцию воды, но отказались от её употребления.
Одна из распространенных гипотез о повышении ошибочных реакций при снижении доминирующей мотивации состоит в нарушении сформированного навыка. Однако нарушение навыка привело бы к одинаковому изменению ошибок.
В данном случае увеличение ошибок при насыщении не является однородным, оно находится в зависимости от результативности предшествующего поведенческого акта — стороны получения предыдущей порции воды. По-видимому, увеличение «неправильных» реакций после насыщения связано не с ошибкой, а с появлением при получении порции воды отрицательной эмоции, с фиксацией её в эмоциональной памяти, и с влиянием в очередном поведенческом акте на выбор цели, направленной на избегание повторного получения этой эмоции.
Для проверки высказанной гипотезы необходимо уточнить возможную фиксацию и активацию в эмоциональной памяти отрицательной эмоции, для чего рассмотрим результаты 2-го эксперимента.
3.2. «Свободный выбор» источников воды при её отсутствии
Второй эксперимент проведен с 8 крысами. Каждый опыт состоял из двух этапов: на первом этапе в течение 30 побежек осуществлялся «Свободный выбор» ИВ без получения порций воды. Второй, завершающий этап проходил по процедуре ПВН для удовлетворения жажды. В течение первых 5 опытов формировалась сигнальная значимость двух различных звуковых стимулов («шум» и «щелчки»): на первом этапе при открытии управляемой защелки включались сигналы «отсутствия воды» (СОВ) без её получения, на втором этапе (процедура ПВН) — сигналы «наличия воды» (СНВ) с её получением. После этой подготовки проведены 6 основных опытов: в 5 опытах на первом этапе по-прежнему включался СОВ, в шестом опыте он заменялся на СНВ.
У всех крыс на первом этапе, несмотря на отсутствие воды, наблюдалась практически незатухающая последовательность побежек к ИВ. Каждая побежка имела характер целенаправленного поведенческого акта, т.к. всякий раз завершалась одной и той же совокупностью консервативных элементов — они подбегали к водной лунке, тщательно вылизывали её сухую поверхность, грызли и царапали её края, отбегали и снова подбегали к ней.
При подаче СОВ все животные проявили тенденцию чередовать выбор источников воды: в среднем по 1200 побежкам (8 крыс, 5 опытов по 30 актов) они в 88±0,8% случаях выбирали противоположный ИВ и только в 12±0,8% повторяли предыдущий выбор (р<0,001).
Объяснение этой закономерности возможно на основе двух предположений: 1) тенденция чередовать поведенческие акты; 2) избегание нерезультативного в предыдущем акте источника воды.
Для их проверки проведен 6-й опыт с включением на первом этапе СНВ. Воду животные по-прежнему не получали, но последовательность выбора резко изменилась. В среднем по 240 побежкам (8 крыс, 1 опыт по 30 актов) частоты смены и повтора выбираемых ИВ составили 51±3,8% и 49,1±3,8% соответственно, с недостоверным их различием для каждого животного и по группе в целом, т.е. эти частоты выровнялись на уровне 50%. Они прекратили избегать тот ИВ, в котором в предыдущем акте получили отрицательный результат. Различия бинарных частот при СОВ и СНВ статистически достоверны (р < 0,001). Если раздражитель сигнализировал возможность достижения положительного результата (при СНВ), влияние предыдущего отрицательного результата на очередной выбор исчезало.
Таким образом, высокая частота выбора противоположного ИВ при включении СОВ связана не с тенденцией чередовать поведенческие акты, а с избеганием того ИВ, в котором перед этим отсутствовала порция воды и была испытана отрицательная эмоция из-за рассогласования в акцепторе действия.
По-видимому, эта эмоция фиксировалась в эмоциональной памяти, а в афферентном синтезе очередного поведенческого акта активировалась и приводила к принятию решения, направленного на избегание её повторного получения. Но при подаче сигнала «наличия воды», предварительно связанного с положительной эмоцией, чередование поведенческих актов исчезало. На стадии афферентного синтеза в памяти активировались следы именно положительных, а не отрицательных эмоций, благодаря чему решения не были направлены на избегание предыдущего нерезультативного ИВ. Это подтверждает высказанную гипотезу об участии эмоциональной памяти в механизмах выбора целенаправленных решений.
В этом эксперименте получена зависимость принятия решения от стороны получения отрицательного результата. Вместе с тем, необходимо рассмотреть особенности «свободного выбора» решений в ситуации с положительными результатами.
3.3. «Свободный выбор» источников воды при многократной переделке стороны её получения
Для этого проведен 3-й эксперимент (по 10 опытов с 5 животными) с многократной переделкой навыка достижения порций воды: сторона их получения в следующем опыте менялась на противоположную. Показатели оценивались в среднем по всем опытам, кроме первого, т.к. в первом опыте переделка ещё отсутствовала.
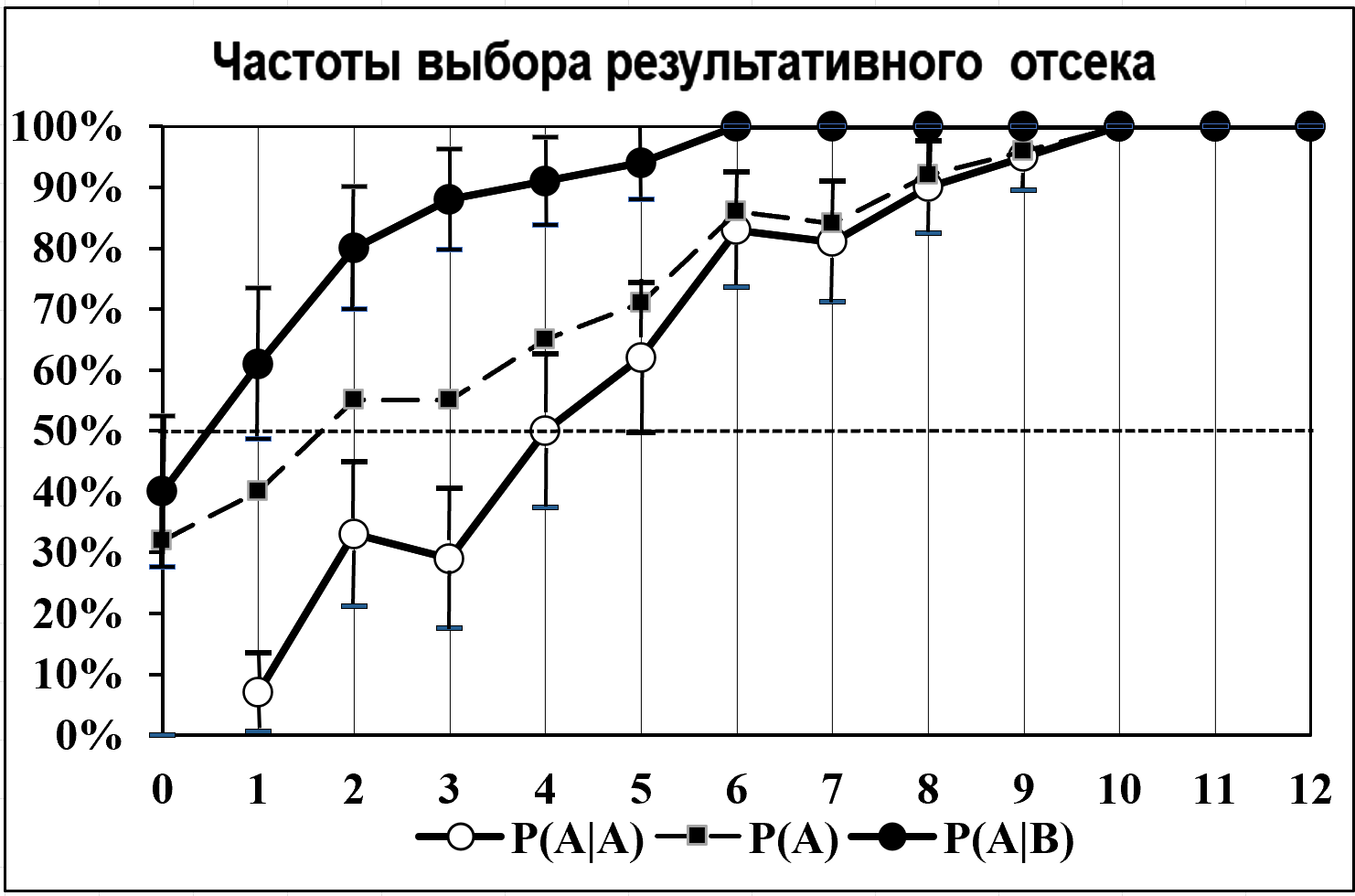
Рисунок 2 - Частоты выбора результативного ИВ лабиринта после каждой порции воды при многократной переделке стороны его получения
До получения первой порции воды (порядковый номер 0 на оси абсцисс) животные выбирали результативный ИВ с «общей частотой» (квадрат) Р(А)=30%. Это низкое значение Р(А) объясняется тем, что этот ИВ был нерезультативным в предыдущем опыте.
Затем, по мере достижений положительного результата, общая частота Р(А) возрастала и после 9-й порции достигла 100%-го уровня. Этот график аналогичен «кривым обучения», известным в экспериментальной и математической психологии , . Он отражает кумулятивное влияние совокупности предшествующих подкреплений на последующее поведение, но без учёта возможной зависимости от предыдущего акта поведения.
Для оценки этой зависимости рассмотрим частоты бинарных комбинаций, представленные на рисунке светлыми и темными кружками.
Видно, что «общие частоты» выбора результативного ИВ Р(А) однонаправленно расщепились на две составляющие, одна из которых отражает условную частоту его повторного выбора Р(А|А), а другая — условную частоту смены побежки, т.е. выбора того же ИВ после предыдущей побежки в противоположный отсек с нерезультативным ИВ Р(A|B).
После 1–4 порции воды Р(A|B) достоверно выше, чем Р(А|А) (Р<0,001); после 5–8 порции различие снизилось (Р<0,01); после 9-й и 10-й порции все частоты выбора результативного ИВ достигли 100%-го уровня. После каждой порции воды значения Р(A|B) выше 50%-го уровня, в чем проявилось избегание повторного выбора нерезультативного ИВ, при достижении которого была испытана отрицательная эмоция. Частоты повторного выбора Р(А|А) после 5-й порции воды превысили 50-й уровень — поведение стало направляться на повторное испытание той положительной эмоции, которая была испытана в предыдущем акте при получении порции воды.
При многократной переделке навыка достижения положительного результата каждый выбор цели определялся не только суммарным числом полученных результатов, но и дифференцированным влиянием на него фиксирующихся в эмоциональной памяти компонентов непосредственно предшествующего результата поведения — положительной или отрицательной эмоции.
4. Обсуждение экспериментальных данных
4.1. Формирование «эмоционально-памятных мотиваций»
Полученные в экспериментах данные подтверждают сделанное предположение о зависимости очередного решения от эмоциональных ощущений в предыдущем поведенческом акте с изменением знака этой зависимости в процессе насыщения. Эта зависимость связана с активацией в эмоциональной памяти следов испытанных эмоциональных переживаний, направляющих поведение на повторное получение прогнозируемой положительной эмоции и на избегание прогнозируемой отрицательной эмоции.
Так как же эмоциональная память «направляет» очередной поведенческий акт на получение положительной и избегание отрицательной эмоции?
По-видимому, на стадии афферентного синтеза активация в памяти следов предыдущих эмоций может приводить к формированию дополнительных мотиваций. Это не доминирующая мотивация, имеющая биологическую природу. Это мотивации психического происхождения, которые можно назвать «эмоционально-памятными мотивациями», направляющими поведение на достижение ранее испытанных положительных, и на избегание ранее испытанных отрицательных эмоций.
На этой основе можно предложить следующую схему эмоционально-мотивированного взаимодействия последовательных актов целенаправленного поведения (рис. 3). В афферентном синтезе очередного поведенческого акта, в зависимости от знака активируемых в памяти следов переживаний, происходит «Формирование эмоционально-памятных мотиваций» достижения ранее испытанной положительной эмоции и избегания ранее испытанной отрицательной эмоции. При их взаимодействии осуществляется принятие очередного решения в виде выбора той цели, которая в большей мере направлена на получение положительного и избегание отрицательного переживания.
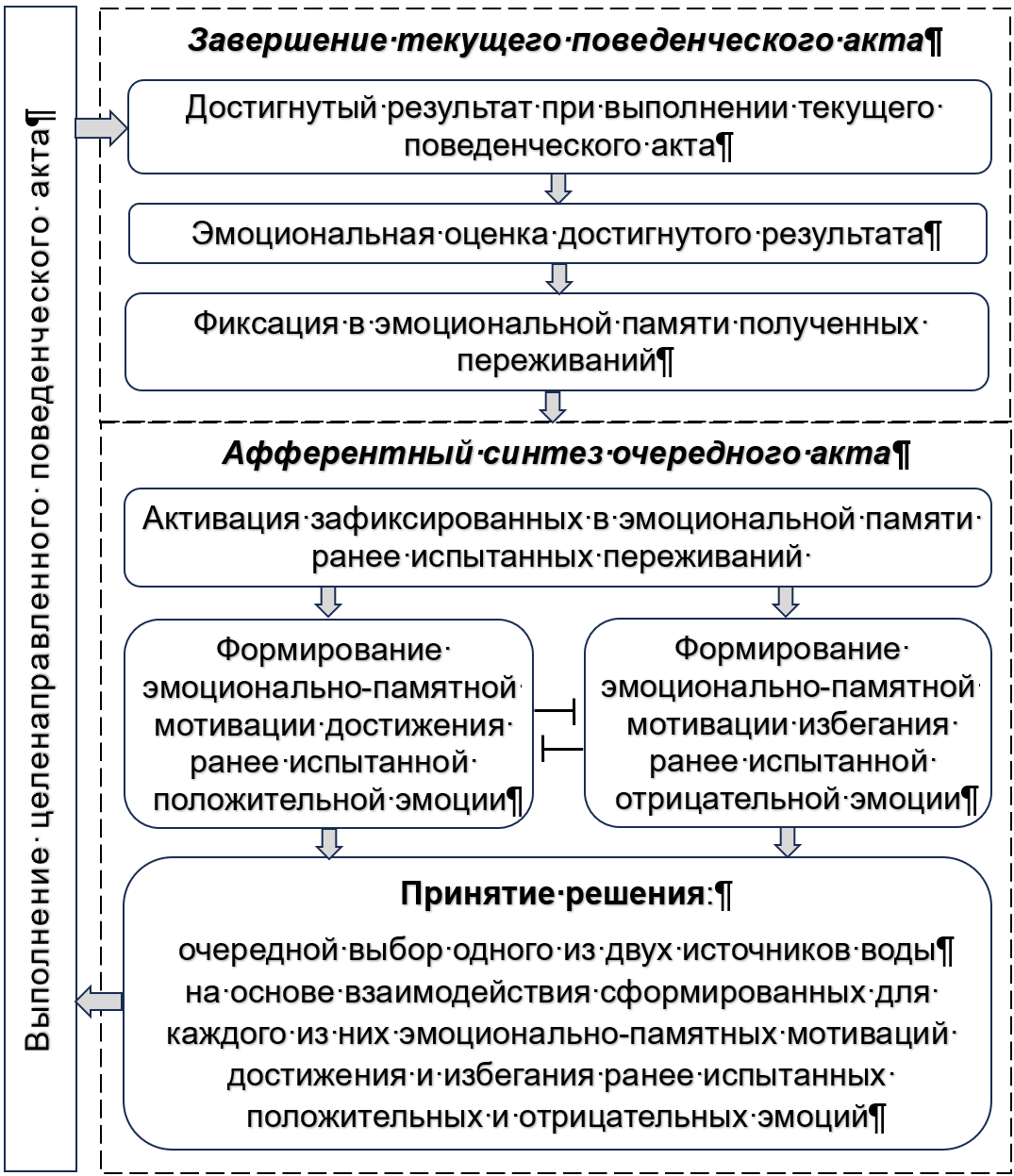
Рисунок 3 - Схема эмоционально-мотивированного взаимодействия последовательных поведенческих актов
Приведенные гипотезы позволяют иначе рассмотреть полученные в первом эксперименте особенности поведения животных после насыщения.
Один из вариантов объяснения наблюдавшихся «холостых реакций» возможен с позиции условного рефлекса. Животному подавался условный сигнал (один из БС), что и вызывало реализацию ранее выработанной инструментальной реакции. Т.е. «автоматический запуск» поведенческих актов, причем с увеличением ошибок. Однако при «автоматическом запуске» число ошибок было бы независимым от результативности предыдущего акта, что противоречит полученным данным о зависимости очередной ошибок от полученного в предыдущем акте результата поведения.
Следует учесть, что при наличии потребности, возникавшие при достижении порций воды позитивные эмоции, могли фиксироваться не только в оперативной, но и в долговременной эмоциональной памяти. Поэтому после насыщения в этой памяти могли активироваться следы испытанных ранее положительных эмоций, что вызывало желание их дальнейшего получения, т.е. формировало «эмоционально-памятную мотивацию достижения положительной эмоции», направленную уже не на удовлетворение потребности, а на получение достигнутых ранее, прогнозируемых удовольствий.
Отсюда следует, что в афферентном синтезе участвует не только доминирующая (биологическая) мотивация, но и «эмоционально-памятная мотивация», формирующаяся под влиянием активируемых в долговременной эмоциональной памяти следов ранее испытанных положительных эмоций.
Животное не только хочет пить, но и стремится к получению тех положительных эмоций, которые ощущало ранее. Именно это и объясняет природу «холостых реакций». При отказах от потребления воды доминирующая мотивация уже отсутствовала. Но обстановочная и пусковая афферентация на стадии афферентного синтеза по-прежнему активировали следы памяти о предыдущих положительных эмоциях, что приводило к появлению «эмоционально-памятной мотивации», под влиянием которой афферентный синтез формировал решение на повторное получение этих положительных эмоций.
В реальной жизни человека так же можно найти случаи появления желаний достижения некоторого результата, даже если никакой потребности в нём нет. Действительно, не только голод заставляет потреблять пищу, но и желание «вкусно поесть». Потребности может не быть, но знакомый запах еды может активировать в памяти следы ранее полученного удовольствия, что формирует желание (т.е. «эмоционально-памятную мотивацию») вновь его получить и направляет на достижение этой цели. Можно привести множество других примеров — табак, алкоголь, наркотики… Жирные и сладкие продукты тоже вызывают положительные эмоции. Поэтому, при отсутствии в организме их потребности, память о ранее полученных удовольствиях может формировать «эмоционально-памятную мотивацию» повторного ощущения этих эмоций, и стремление к потреблению этих продуктов.
Если «холостые реакции» действительно вызывались «эмоционально-памятной мотивацией достижения», то что же приводило через множество побежек к полному их прекращению?
Процесс насыщения имеет две стороны. Одна из них, которая чаще другой принимается во внимание, заключается в уменьшении соответствующей потребности.
Другая сторона этого процесса состоит в изменении биологической оценки самого результата, в инверсии биологического значения порций воды с положительного на отрицательное, в появлении отрицательных эмоций при достижении ставшего ненужным результата, в накоплении в долговременной эмоциональной памяти следов отрицательных переживаний и в их влиянии на принятие очередного решения. Активация накапливающихся следов негативных переживаний могло приводить к формированию еще одной «эмоционально-памятной мотивации» — мотивации избегания испытанных ранее при получении воды отрицательных эмоций.
После насыщения, во время «холостых реакций», происходила «борьба» двух «эмоционально-памятных мотиваций». Животные прекращали поведение и засыпали не столько от усталости, сколько от подавления «эмоционально-памятной мотивации достижения положительных эмоций» усиливающейся «эмоционально-памятной мотивацией избегания отрицательных переживаний».
4.3. Структура мотиваций целенаправленного поведения до и после насыщения
Можно предположить следующую структуру формирования мотиваций до и после насыщения (рис. 4).
У животного с опытом удовлетворения потребности действующая мотивация состоит из трёх частей: биологическая (жажда) и, по крайней мере, две «эмоционально-памятные мотивации», направленные на достижение и избегание ранее испытанных эмоций.
Источником всех мотиваций является «Доминирующая мотивация, вызванная потребностью» и действующая на стадии «До насыщения». При этом активация в эмоциональной памяти следов позитивных эмоциональных переживаний приводит к прогнозу их повторного получения и, тем самым, к появлению «Эмоционально-памятной мотивации достижения ранее испытанных положительных эмоций».
До насыщения «эмоционально-памятная мотивация достижения» действует совместно с доминирующей мотивацией, усиливая ее. Но эта мотивация продолжает действовать и «после насыщения». Это — тоже целенаправленное поведение, в котором цель состоит в достижении прогнозируемых позитивных ощущений.
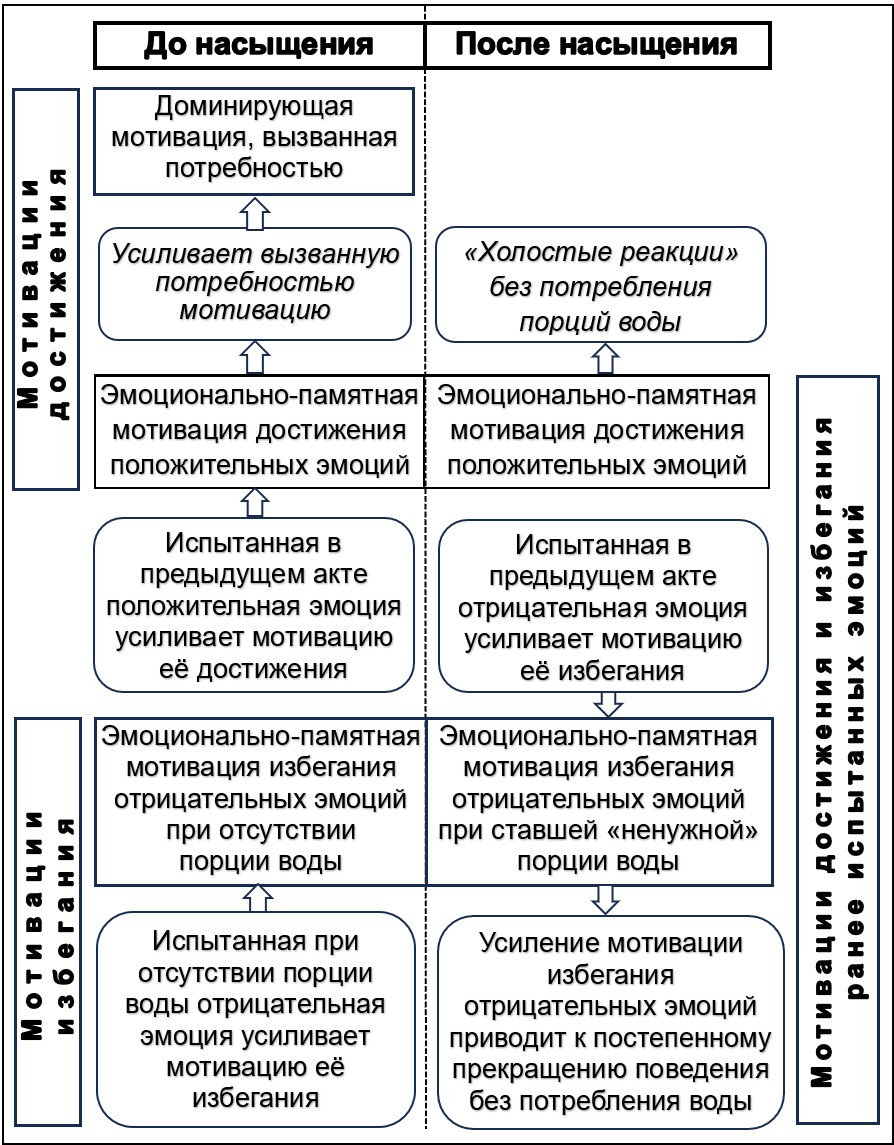
Рисунок 4 - Структура мотиваций достижения и избегания до и после насыщения
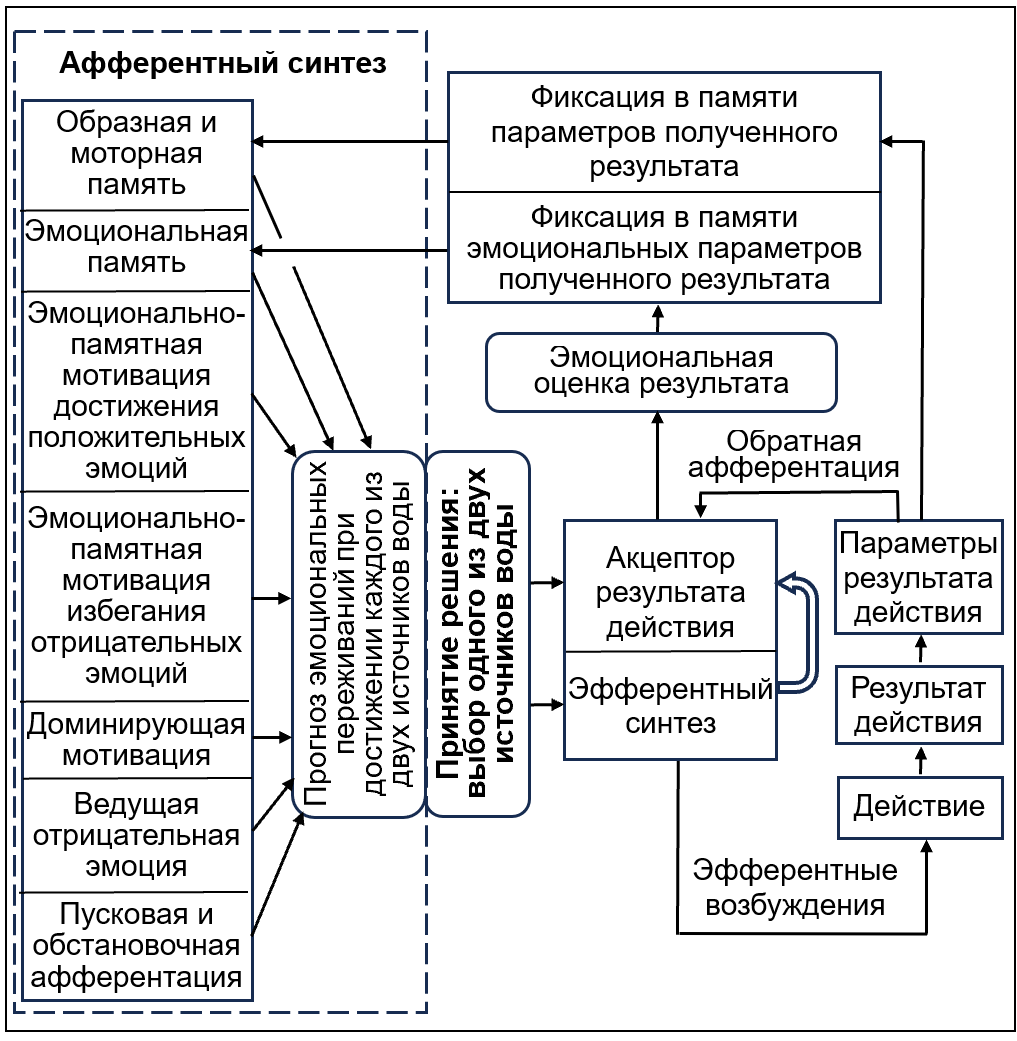
Рисунок 5 - Модифицированная модель функциональной системы целенаправленного поведения в ситуации последовательного выбора
Целью решения является достижение положительных и избегание отрицательных эмоций, следы которых активированы в долговременной памяти. На этой основе осуществляется «Эфферентный синтез» и формирование «Акцептора результата действия». Далее совершается целенаправленное «Действие» и получение «Результата действия» с его «Параметрами».
Согласно П.К. Анохину , , сравнение параметров полученного результата с акцептором (с предвосхищением) результата выполняемого действия приводит к положительным или к отрицательным эмоциям, т.е. к «Эмоциональной оценке результата». Поэтому, совместно с «Фиксацией в памяти параметров полученного результата», в «Эмоциональной памяти» осуществляется фиксация его «эмоциональных параметров», т.е. внутренних переживаний.
Благодаря этому в афферентном синтезе очередного поведенческого акта участвует дополнительный компонент — «Эмоциональная память» о ранее испытанных эмоциональных переживаниях. Этот компонент не является текущей эмоцией, это — воспоминание о тех эмоциональных ощущениях, которые были испытаны при достижении предыдущих результатов. При активации в памяти ранее полученных эмоций в афферентном синтезе происходит формирование ещё двух амбивалентных компонентов — «Эмоционально-памятной мотивации достижения» и «Эмоционально-памятной мотивации избегания» ранее испытанных положительных и отрицательных эмоций. Именно эти два компонента определяют целенаправленное поведение на всех этапах снижения доминирующей мотивации — до и после насыщения.
Таким образом, в афферентном синтезе поведенческого акта участвует не только память о внешних событиях, но и память о ранее испытанных внутренних (эмоциональных) переживаниях, которые приводят к формированию дополнительных «эмоционально-памятных мотиваций», связанных не с потребностью, а с ранее пережитыми эмоциями.
Безусловно, главную роль играет «Мотивация, вызванная потребностью». Именно она в первую очередь определяет нацеленность поведения на достижение необходимого результата. Но независимо от неё, и даже после её удовлетворения, поведение направляется дополнительными компонентами афферентного синтеза — «эмоционально-памятными мотивациями» достижения положительных и избегания отрицательных эмоций.
По-видимому, в целенаправленном поведении все мотивации — это работа с эмоциями: устранение возникшей при наличии потребности «Ведущей отрицательной эмоции», а также «эмоционально-памятные мотивации достижения» и «эмоционально-памятные мотивации избегания» прогнозируемых эмоциональных переживаний.
5. Заключение
Закономерности последовательного выбора крысами целей поведения выявили зависимость принимаемых решений от результативности предыдущего поведенческого акта с различным знаком этой зависимости до и после насыщения. При полноценной жажде зависимость проявилась в виде избегания неуспешного и повторного выбора успешного поведенческого акта.
После насыщения наблюдались «холостые реакции» животных с отсутствием потребления порций воды. При этом зависимость от результативности проявилась в избегании повторного выбора изменившего биологическое значение «положительного результата».
В основе этих зависимостей, скорее всего, находится эмоциональная память о полученных ранее положительных или отрицательных переживаниях и их активация на стадии афферентного синтеза очередного поведенческого акта. Активация в памяти этих следов приводит к формированию «эмоционально-памятных мотиваций» достижения и избегания возникших в предыдущих актах положительных и отрицательных эмоций соответственно. Это не доминирующая мотивация, имеющая биологическую природу. Это мотивации психического происхождения, которые «направляют» поведение на достижение ранее испытанных положительных, и на избегание ранее испытанных отрицательных эмоций.
«Холостые реакции» животных после насыщения — это тоже целенаправленное поведение, направляемое «эмоционально-памятной мотивацией» достижения ранее испытанных положительных эмоций. Цель этого поведения состоит не в получении порций воды, а в достижении ранее испытанных позитивных эмоциональных ощущений.
Биологическое значение порций воды в процессе насыщения изменяется с позитивного на негативное, приобретая черты отрицательно-эмоционального характера. В результате этого происходит перестройка целенаправленного поведения, связанная с появлением избегания испытанной в предыдущем акте отрицательной эмоции при получении, ставшего отрицательным, «положительного результата». Прекращение поведения и засыпание животных после насыщения происходит при балансе амбивалентных «эмоционально-памятных мотиваций», наступающим из-за накопления в эмоциональной памяти следов отрицательных эмоций и усиления «эмоционально-памятной мотивации» их избегания при достижениях результата, биологическое значение которого изменилось на негативное.
С учетом этого предложена схема эмоционально-мотивированного взаимодействия последовательных поведенческих актов. В афферентном синтезе очередного акта поведения, наряду с доминирующей мотивацией, участвуют «эмоционально-памятная мотивация достижения» ранее испытанной положительной эмоции и одновременно «эмоционально-памятная мотивация избегания» ранее испытанной отрицательной эмоции, что находится в зависимости от знака активируемых в эмоциональной памяти следов переживаний.
На основе их взаимодействия осуществляется принятие очередного решения в виде выбора того поведенческого акта, который в большей мере направлен на получение положительной и избегание отрицательной эмоции.
Предложена структура мотиваций до и после насыщения. До насыщения совместно с доминирующей мотивацией, усиливая ее, действует «эмоционально-памятная мотивация достижения». Но эта мотивация продолжает действовать и после насыщения без доминирующей мотивации, что и приводит к «холостым реакциям». Кроме того, после насыщения формируется «эмоционально-памятная мотивация избегания ранее испытанных отрицательных эмоций» при достижении, ставшего «ненужным», результата, приводящая к постепенному прекращению поведенческих актов.
На этой основе проведена модификация схемы функциональной системы последовательного выбора целей поведения. В афферентном синтезе очередного поведенческого акта участвует не только память о внешних событиях, но и дополнительный компонент — «Эмоциональная память» о ранее испытанных внутренних переживаниях. Этот компонент не является эмоцией, это — активация и прогнозирование тех эмоциональных ощущений, которые были испытаны при достижении предыдущих результатов. Именно это приводит к формированию дополнительных «эмоционально-памятных мотиваций», связанных не с потребностью, а с ранее пережитыми эмоциями.
Эмоциональная память и эти две мотивации определяют перестройку целенаправленного поведения животных на разных стадиях насыщения.